2025 年 9 月 15 日,Min Lab(团队详情在文末)在《Advanced Science》期刊上发表了一篇题为“SpaBatch: Deep Learning-Based Cross-Slice Integration and 3D Spatial Domain Identification in Spatial Transcriptomics”的研究论文,其中指出随着来自不同组织、个体及技术平台的空间转录组学(ST)数据迅速积累,迫切需要一种稳健可靠的多切片整合框架,以实现三维空间结构域的识别。然而,现有方法主要集中于单一切片内的二维空间结构域识别,难以充分考虑切片间的空间关联性以及批次效应的校正,从而限制了跨切片三维空间结构域识别的准确性。本研究提出SpaBatch,一种用于整合与分析多切片ST数据的新型框架,可有效校正批次效应,并实现跨切片的三维空间结构域识别。为展示SpaBatch的优势,作者将其应用于八组真实ST数据集,包括来自不同个体的人类皮层切片、采用两种不同技术生成的小鼠脑切片、小鼠胚胎切片、人类胚胎心脏切片、HER2+乳腺癌组织以及基于MERFISH平台检测的小鼠下丘脑切片。全面验证结果表明,SpaBatch在三维空间结构域识别方面 consistently 优于当前先进方法,同时有效校正了批次效应。此外,SpaBatch能够高效捕捉切片间保守的组织结构与癌症相关亚结构,并利用有限的标注信息预测未标注切片中的空间结构域,凸显其在组织结构解析与发育生物学研究中的潜在应用价值(文章较长(可到总结-概述-分析方法最后部分)文章数据分析复现代码实例在文末)。
注:本文涉及的公式及函数可以直接复制公式到LaTeX编辑器查看具体公式,我这边推荐:(https://arachnoid.com/latex/或 https://www.quicklatex.com/)🤓🤓
简介
空间转录组学(ST)是一项快速发展的技术,能够在保留组织空间位置信息的同时获取基因表达数据,为解析细胞的空间组织模式及组织结构的构建提供了关键见解。近年来,多种具有不同空间分辨率的ST技术相继问世。例如,10x Visium采用直径为55微米的捕获探针,在“点”(spot)水平上定量转录本丰度,每个点通常覆盖多个细胞。Slide-seq及其改进版本Slide-seqV2的空间分辨率约为10微米,接近单细胞水平。Stereo-seq则实现了500纳米(0.5微米)的空间分辨率,在空间转录组技术中实现了突破,兼具超高清分辨率与厘米级组织覆盖能力。
在ST数据分析中,空间结构域识别是核心任务之一,旨在将组织切片划分为多个功能区域,确保同一结构域内的空间点具有相似的基因表达模式。近年来,针对单一切片的二维空间结构域识别已取得显著进展。通过图神经网络整合空间信息的方法,以及将空间转录组数据与组织病理图像结合的深度学习策略,显著提升了单切片ST数据分析中空间结构域识别的准确性。然而,生物体与组织的生物学活动本质上是三维的。随着在不同条件、技术与平台下生成的ST数据持续积累,迫切需要对多个组织切片的ST数据进行联合分析,以揭示复杂的组织架构并解析跨个体的生物学规律。然而,多切片ST数据的整合与三维空间结构域识别仍面临诸多挑战,如空间错位与批次效应。已有若干方法尝试解决这些问题,例如,某方法在初始阶段进行多切片坐标对齐,并利用PASTE算法对潜在嵌入空间中的数据进行批次效应校正,但其准确性高度依赖于空间坐标对齐的精度。另一方法采用深度图神经网络进行表征学习,并引入领域对抗网络(DAN)以校正批次效应,但其训练过程耗时较长,且未能实现不同样本间更有效的整合。还有一种方法利用STAGATE对多切片联合数据进行表征学习,并基于互近邻(MNN)策略调整潜在嵌入空间,旨在缩小正样本对之间的距离,同时扩大负样本对之间的距离。然而,该方法可能忽略来自相同空间结构域的非互近邻配对,同时错误地将属于不同结构域的互近邻配对纳入考虑。另有方法利用深度图神经网络进行表征学习,并对潜在嵌入进行预聚类,以定义跨样本的正负样本对,进而在训练过程中采用三元组对比学习策略,逐步拉近正样本对、推远负样本对,从而校正批次效应。然而,预聚类过程可能导致聚类分配错误,进而对后续训练产生不利影响。
此外,诸如stlearn和STitch3D等方法需要整合来自相同组织的单细胞RNA测序数据或组织学图像等额外数据源,这限制了其广泛应用。现有方法在批次效应校正方面往往能力不足,且在跨切片空间结构域识别中表现出不稳定性。同时,大多数方法无法输出经过校正的基因表达数据,从而限制了其在下游分析中的应用价值。为克服上述局限,作者提出一种新型多切片整合分析框架——SpaBatch(图1)。具体而言,SpaBatch首先采用掩码策略对输入的基因表达谱进行扰动,以增强模型对空间转录组数据中常见的缺失值和技术噪声的鲁棒性。在训练过程中,SpaBatch采用预训练与微调相结合的双阶段策略。在预训练阶段,利用变分图自编码器(VGAE)联合建模基因表达与空间邻接信息,学习能够反映潜在空间转录组格局的低维隐含嵌入。在微调阶段,引入深度嵌入聚类(DEC)模块,迭代优化嵌入空间,增强识别出的结构域内部的空间紧凑性。为进一步应对切片间的异质性并减轻批次效应,SpaBatch引入基于读出机制的三元组对比学习策略,该机制可跨切片聚合多个正样本,同时最大化正负样本对之间的距离。双阶段微调的引入使模型能够在切片间对齐生物学上一致的结构,同时保留实现精确三维空间结构域识别所需的判别性空间特征。
图1. SpaBatch的架构示意图。A)输入数据与数据预处理。B)SpaBatch的主干网络。C)下游分析。图A(输入数据,左侧)中的脑切片示意图。作者将SpaBatch应用于八组来自不同组织类型(人背外侧前额叶皮层、小鼠脑、人胚胎心脏、HER2+乳腺癌)、物种及技术平台(10x Visium、ST、Stereo-seq、MERFISH)的空间转录组数据。在DLPFC数据集上的实验结果表明,SpaBatch在空间结构域识别(DLPFC数据集样本3上的平均ARI为0.613,而其他所有方法均低于0.6)和批次效应校正(相较于STG3Net和DeepST等方法,SpaBatch实现了更优的平衡性能)方面均优于当前先进方法。在小鼠脑数据集中,SpaBatch进一步整合了来自两种不同空间转录组平台的数据,从宏观与微观层面全面解析了复杂的组织空间结构——例如三突触海马回路,并通过标志基因表达进行了验证。在包含多个发育阶段(受孕后4.5至6.5周)切片的人胚胎心脏数据集中,SpaBatch有效捕捉了心脏发育过程中的动态时空变化。有趣的是,在整合HER2+乳腺癌数据集时,SpaBatch采用半监督学习策略,利用一个已标注切片的有限信息,在其他未标注切片中成功识别出癌变与非癌变区域。SpaBatch在该任务中也表现出稳健的聚类性能(中位ARI为0.361,较次优基线方法提升38%),展示了其在病理学研究中的应用潜力。下游分析识别出具有空间特异性表达的基因(如DLPFC数据集中皮层表达的TMSB10,以及HER2+乳腺癌肿瘤区域表达的ERBB2),并通过基因本体(GO)富集分析揭示了组织特异性的生物学过程(如肿瘤微环境中的免疫应答)。总体而言,SpaBatch为跨多切片的空间转录组数据整合提供了一个灵活而有效的工具,能够实现批次效应校正,并提升三维空间结构域识别的准确性与稳定性。
结果概况
SpaBatch在DLPFC数据集中有效校正批次效应并精确识别空间结构域
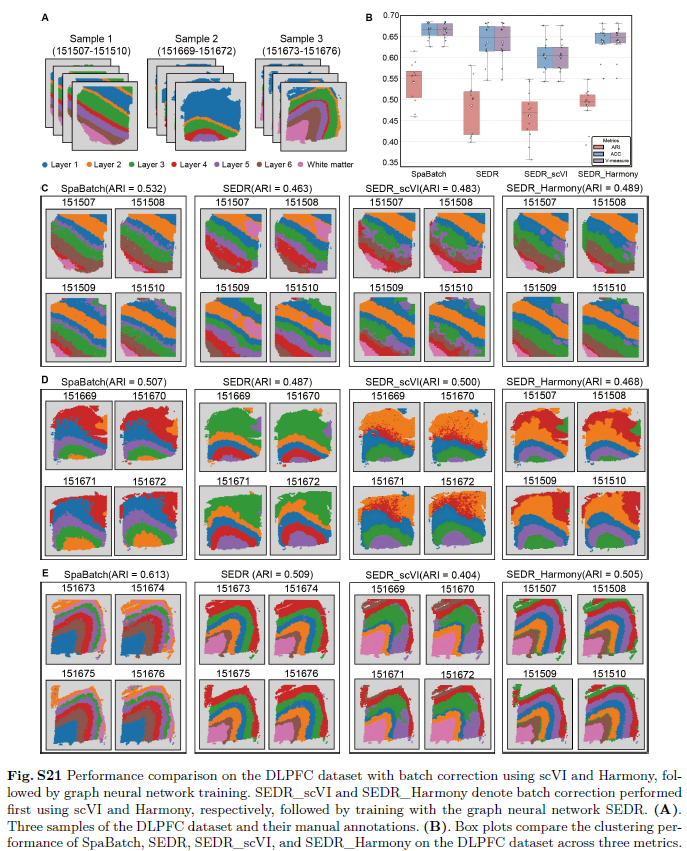
为定量评估SpaBatch在多切片联合分析中的空间结构域识别与批次效应校正能力,作者首先将其应用于10x Genomics Visium平台测得的人背外侧前额叶皮层(DLPFC)数据集。作者将SpaBatch与六种当前先进方法进行比较,采用ARI、ACC和V-Measure等指标评估其在空间结构域识别方面的性能,同时利用iLISI和cLISI指标评估其在批次效应校正和多切片整合效果上的表现。作者将来自不同供体的样本分为三组,每组样本包含四个相邻的组织切片(图2A)。首先在样本3上进行评估,发现SpaBatch、STAligner和STG3Net识别出的空间结构域在相同皮层内混合良好,而不同皮层则按照从第1层到第6层及白质层(WM)的空间结构有序排列。与后两种方法相比,SpaBatch识别出的边界更清晰、形态更精确。其他方法在第1至第4层内的空间结构域识别存在明显偏差。在聚类准确性方面,SpaBatch的ARI值最高,平均达到0.613,优于STAligner(ARI = 0.578)和STG3Net(ARI = 0.576),并显著优于其余方法(图2C)。作者在样本1和样本2上进行了相同实验,结果表明SpaBatch在三维空间结构域识别方面同样优于其他方法(图S3,支持信息)。分别对样本1至3的全部12个切片进行实验并计算ARI值(表S4,支持信息),结果显示SpaBatch的中位数和平均ARI得分均最高,进一步凸显其在DLPFC数据集上空间结构域识别的优越性(图2B;图S4A,支持信息)。此外,作者通过比较不同随机种子下的聚类准确性,进一步验证了SpaBatch的稳健性,发现其对随机种子的变化不敏感(图S4B,支持信息)。
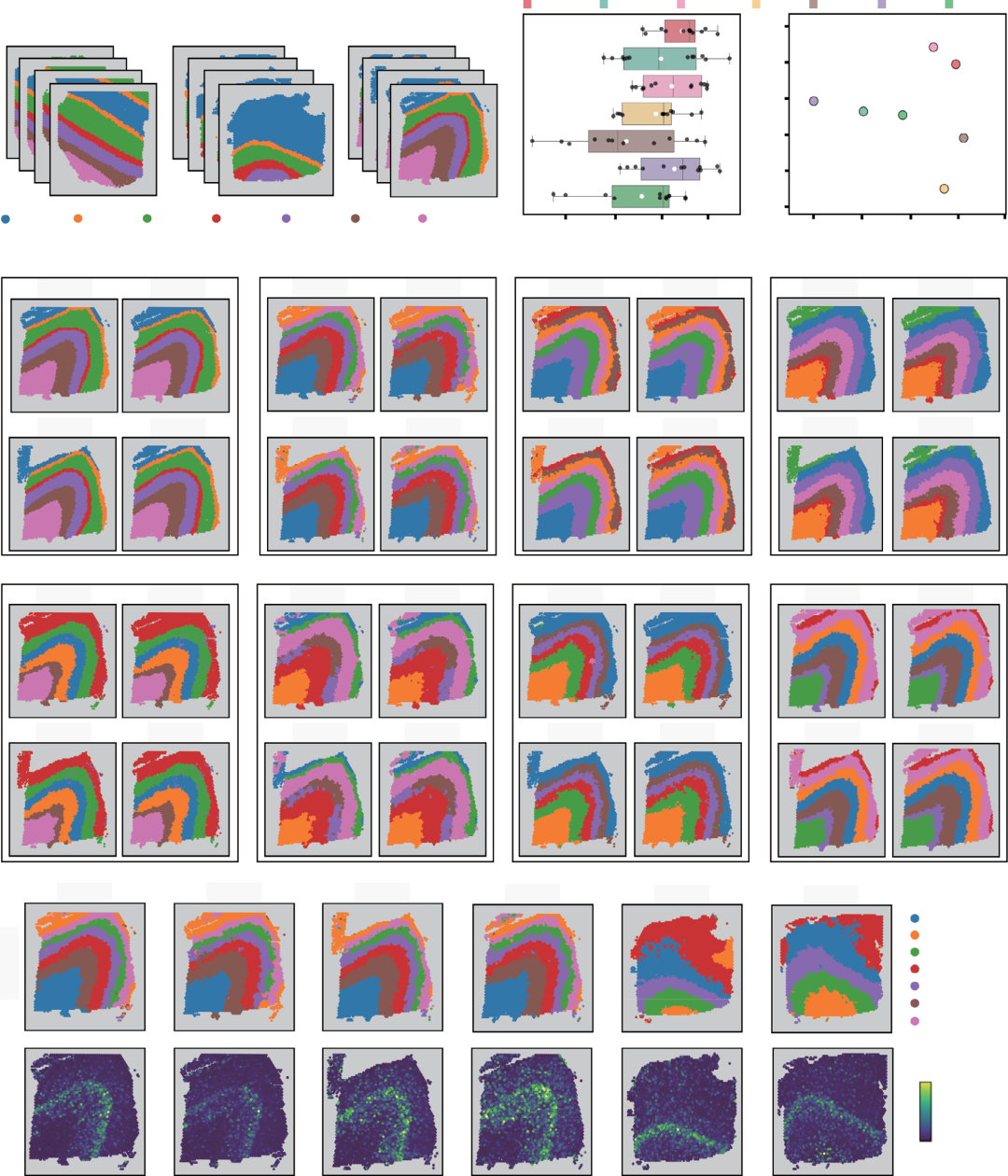
图2. DLPFC数据集多切片联合分析结果。A)DLPFC数据集的三个样本及其人工注释。B)SpaBatch与其他方法在DLPFC数据集三个样本共12个切片上计算的ARI值箱线图。箱线图中,中间线和白色实心点分别代表中位数和均值。左侧的蜂群图展示了所有切片上的准确性分布。右侧显示了SpaBatch与其他方法在DLPFC数据集三个样本上计算的iLISI和cLISI得分,横轴表示批次混合得分,纵轴表示空间结构域混合得分,越靠近右上角的点表示性能越优。C)SpaBatch与基线方法对DLPFC数据集样本3的四个切片进行整合与空间结构域识别的结果。D)SpaBatch识别出的六个切片的空间结构域(上图),以及第5层标志基因TMSB10的空间表达模式(下图)。
Scanpy生成的均匀流形近似与投影(UMAP)可视化结果显示,在UMAP空间中不同批次之间无明显分离,表明DLPFC数据集中的批次效应本身不显著(图3)。然而,与基于深度学习的模型相比,Scanpy的聚类边界相对模糊,聚类结果的空间紧凑性和空间连续性较差,凸显了传统方法在处理复杂空间转录组模式上的局限性。其他深度学习方法均生成了结构清晰、与人工注释高度吻合的皮层分层结构。其中,SpaBatch在精确捕捉空间结构域方面表现尤为突出,其UMAP可视化结果与人工标注的皮层分布高度一致。此外,SpaBatch在不同批次间实现了平滑且均匀的混合,这一特性在跨切片的批次整合中得到了充分体现。值得注意的是,尽管部分深度学习模型(如STAligner和SpaGIC)在一定程度上提升了聚类紧凑性,但其经过批次校正后的UMAP可视化仍表现出不同批次样本再次分离的趋势,这种“过度校正”现象提示这些模型在整合过程中可能过度强调了批次效应的消除。iLISI和cLISI的数值也间接反映了SpaBatch在实现有效切片混合与批次效应校正方面的优势。
图3 样本3(切片151673-151676)嵌入结果的UMAP可视化,分别按识别出的结构域(左)、真实情况(中)和批次校正后(右)进行着色。
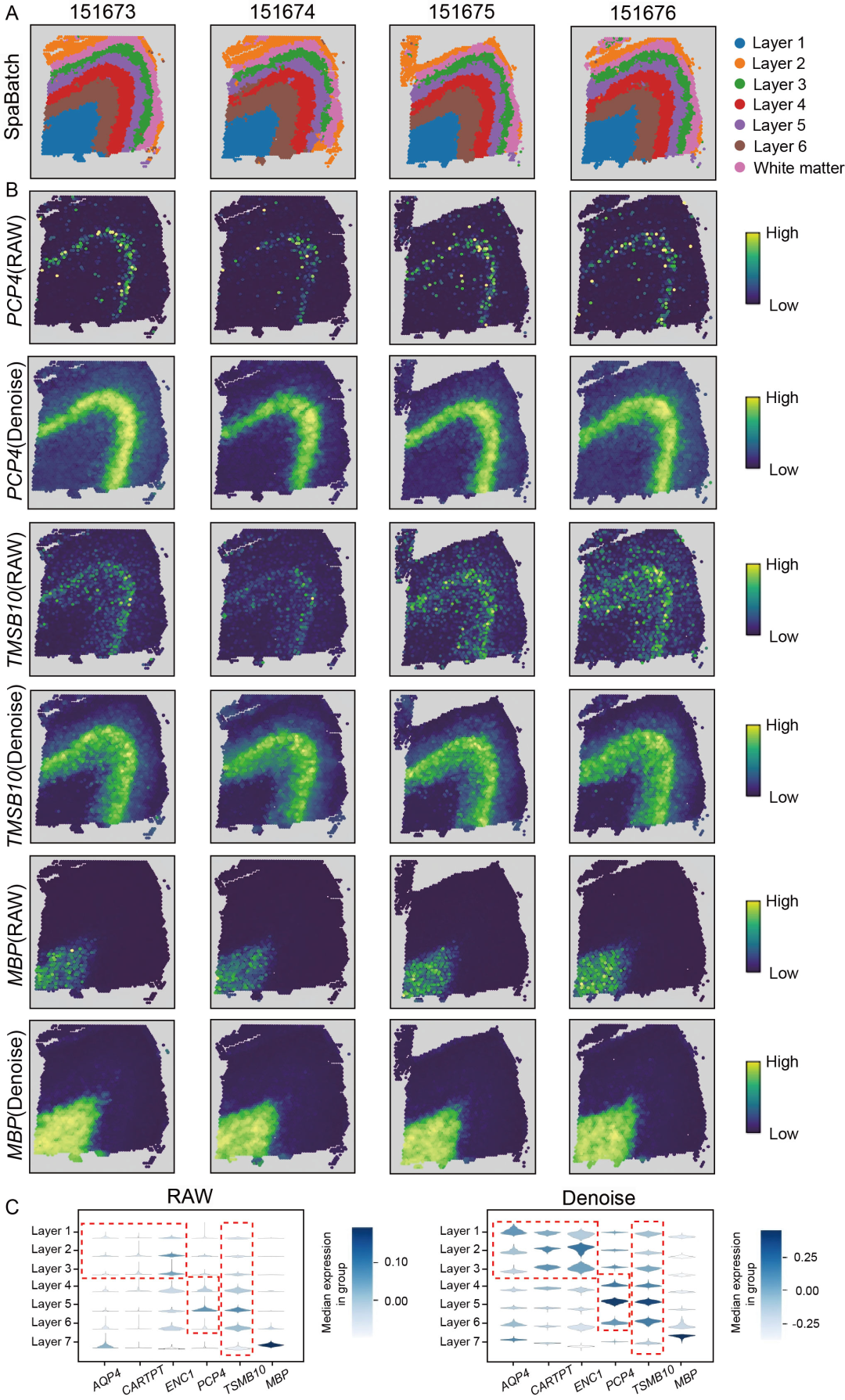
尽管STG3Net较高的cLISI得分体现了其在批次效应校正上的优势,但其iLISI得分较低,提示可能损失了部分生物学信号;而DeepST较高的iLISI得分反映了其对生物学信号的良好保留,但cLISI得分偏低,表明其批次效应校正不足。SpaBatch则表现出均衡的性能,在cLISI上表现优异的同时保持了较高的iLISI得分,体现了其在有效校正批次效应的同时保留关键生物学信号的能力(图2B右侧)。SpaBatch不仅在空间结构域识别上实现了更高准确性,也确保了跨批次整合的高度一致性与连贯性。这一优势在其他样本中同样得以保持(图S5和S6,支持信息)。作者通过差异表达分析并结合已有研究,进一步鉴定了各皮层的标志基因,例如AQP4(第1层)、CARTPT(第2层)、ENC1(第3层)、PCP4(第4层)、TMSB10(第4层和第5层)以及MBP(白质层),这些基因在不同皮层间表现出显著的表达差异(图S7)。通过对第5层标志基因TMSB10进行可视化,作者展示了SpaBatch能够有效识别标志基因,并描绘出不同样本间共享的组织结构模式(图2D)。
SpaBatch全面描绘小鼠脑部从宏观到微观的结构
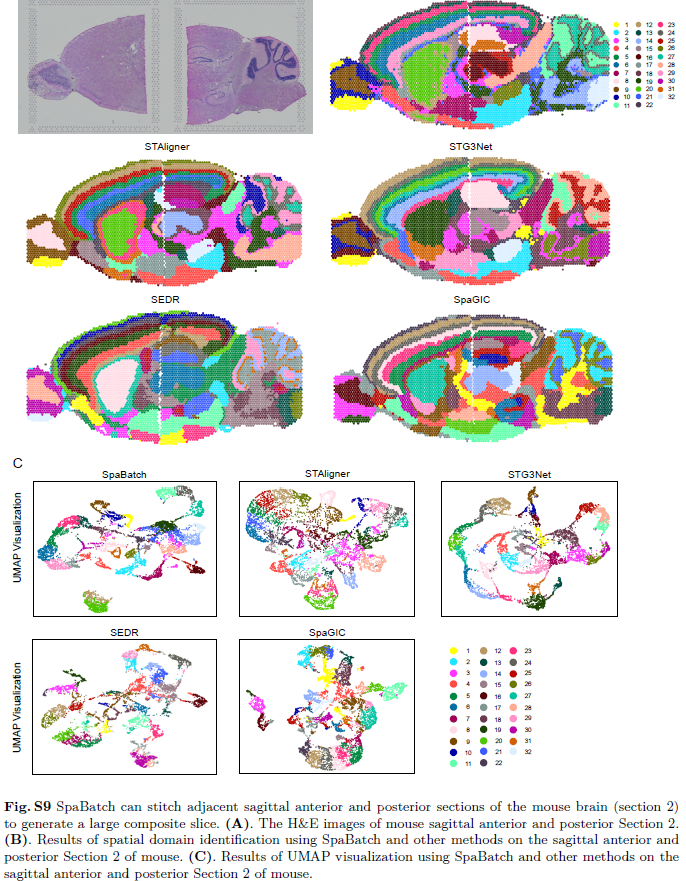
接下来,作者在更复杂的组织切片上评估了SpaBatch的表现。作者使用10x Visium协议生成的小鼠脑矢状面数据的第一和第二切片进行了实验(图4A;图S8A和S9A,支持信息)。第一和第二切片均包含前脑和后脑配对样本。作者将第一和第二切片的前脑与后脑区域分别进行整合处理。在这两个数据集中,只有第一切片的前脑区域有人工注释,作者以此作为基准(图S8B,支持信息)。这些数据为理解小鼠脑部的整体解剖结构提供了全局视角。
图4 SpaBatch从宏观层面全面描绘矢状面小鼠脑结构。A)第一切片前脑和后脑矢状面的H&E染色图像及其对应的特异性空间亚结构域。B)来自Allen小鼠脑参考图谱的第一切片前脑矢状面人工注释。C)SpaBatch与其他方法对第一切片小鼠脑矢状面进行整合后得到的空间结构域识别结果。作者使用红色方框和数字突出显示特定的空间亚结构域。D)比较SpaBatch与其他方法获得的ARI值的柱状图。E)SpaBatch识别出六个不同的精细空间亚结构域(上图),以及与这些亚结构域相关的标志基因的空间表达模式(下图)。
首先,作者测试了SpaBatch、STAligner、STG3Net、SEDR和SpaGIC在第一切片上的整合能力,并将所有方法的聚类数量设置为32以作比较。Allen脑图谱提供的小鼠脑图谱被用作参考(图4B)。研究发现,SpaBatch、STAligner、STG3Net和SEDR均能够识别第一和第二切片连续切片相邻区域之间的共享簇,主要包括皮层、海马体、丘脑和下丘脑。对于更为精细的组织结构,SpaBatch展现了更强的检测能力。前脑的主要嗅球以及后脑的小脑皮层(图4C中的红色方框1和4)由SpaBatch检测出的结果与Allen脑图谱的小鼠皮层参考图谱高度一致。虽然SEDR同时识别了这两个空间结构域,但其在小脑皮层区域的准确性仍不及SpaBatch。此外,作者进一步检查了位于后脑的海马体背侧和腹侧区域(图4C中的红色方框2和3),这些区域跨越前后脑,增加了空间结构域连续识别的难度。背侧海马区包括CA1、CA2、CA3及齿状回(DG),共同构成了经典的三突触环路,这对记忆形成和检索至关重要。在所有方法中,仅SpaBatch准确识别了海马体的腹侧区域并完全认出了由CA1、CA2、CA3组成的阿蒙角以及齿状回(DG)。其他方法未能在此区域内实现准确识别。最后,对于两个切片共有的皮层区域,SpaBatch能够全面识别并精确区分不同的亚区域,包括分子层(第1层)、外颗粒细胞和锥体层(第2/3层)、内颗粒层(第4层)、内锥体层(第5层)和多形层(第6a层和第6b层),这些区域与定义的空间结构域15、16、18、19、25和28相吻合(图4C)。在第一切片前脑部分的ARI得分方面,SpaBatch取得了最高的性能(ARI = 0.44),优于其他所有方法(图4D)。此外,SpaBatch通过估算基因表达增强了结构域标志基因的信号。例如,Neurod6基因的表达与第11结构域中识别出的CA区域相符,而Vxn和Lamp5则对应于代表第2/3层和第6a、6b层的结构域15和16。此外,Dner和Shf的组合完整地代表了在结构域19和29中识别的小脑区域(图4E;图S8C,D,支持信息)。这一例子进一步突显了SpaBatch在减轻批次效应的同时提供小鼠脑矢状面全面描绘的能力,这一优势在第二切片中也得到了体现(图S9,支持信息)。
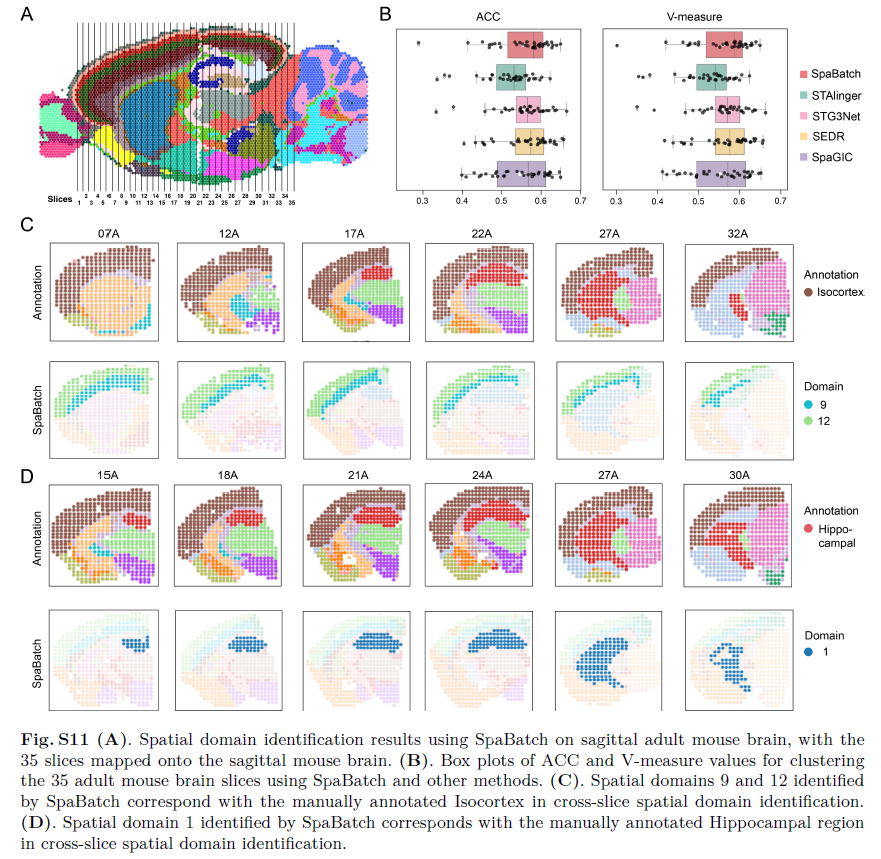
为进一步从微观层面探索成年小鼠脑部结构,作者收集了沿前-后(AP)轴分布的35个冠状切片,其中包含15种聚类类型的注释信息。这些切片覆盖了从嗅球到小脑出现的整个小鼠脑区(图S10,支持信息)。基于作者的聚类结果和人工注释,作者将这些冠状切片的位置映射到Allen脑图谱参考图以及SpaBatch获得的矢状切片上(图5A;图S11,支持信息),从而更全面地理解不同解剖平面上脑区的空间分布与组织架构。通过分析这些切片,作者可以从更精细的微观视角观察成年小鼠脑在不同前-后位置的渐进性变化。该多切片联合分析任务具有挑战性,要求方法能够处理数十个切片间的批次效应,并区分成年小鼠脑内空间结构域的细微差异。
图5 SpaBatch从微观层面全面描绘矢状面小鼠脑结构。A)基于SpaBatch识别的空间结构域结果和人工注释,将35个切片的位置映射到Allen脑图谱参考图上。B)SpaBatch与其他方法在35个冠状面小鼠脑切片上计算的ARI值箱线图(左图);以及在该数据集上计算的iLISI和cLISI得分(右图)。C–F)SpaBatch沿前-后(AP)轴在不同冠状切片中准确识别空间结构域,并通过Allen脑图谱参考对应到特定组织结构。SpaBatch还能进一步识别出人工注释中未划分的区域。G)SpaBatch识别的35个切片中各空间结构域比例的堆叠柱状图。H)海马体的结构域标志基因Neurod6,以及皮层的结构域标志基因Lamp5和Ncald,在矢状面和冠状面小鼠脑切片中均表现出一致的空间表达模式。
SpaBatch在35个切片中识别出的空间结构域表现出强连续性(图S11C,D,支持信息),且所有识别结果均在Allen脑图谱参考中得到验证。在最早的成年小鼠脑切片(01A-03A)中,主要区域包括新皮层和嗅觉区。SpaBatch不仅准确识别了这两个区域,还检测出人工注释中未区分的亚区,特别是将新皮层精确划分为FRP 1和FRP 2/3(图5C)。随着海马体逐渐扩展(16A-20A),SpaBatch准确捕捉到四个关键空间结构域,与人工注释高度一致(图5D)。当海马体继续扩展、下丘脑逐渐缩小、中脑开始显现时(22A-26A),这些动态变化也被SpaBatch清晰识别(图5E)。当海马体过渡为后海马区(33A-35A)时,SpaBatch将该区域进一步划分为内嗅皮层(ENT)、下托(SUB)和前下托(PRE)(图5F),这些亚区在原始注释中并未区分,而它们在空间感知、记忆形成以及皮层与海马间信息传递中发挥关键作用。通过对全部35个冠状切片的聚类比例进行分析,SpaBatch清晰呈现了小鼠脑组织组成及其动态变化过程。例如,海马结构域在15A切片出现,在31A切片后消失(图5G)。值得注意的是,在矢状切片中,SpaBatch识别出多个具有空间特异性的标志基因,包括Neurod6(海马体)、Lamp5(皮层)和Ncald(皮层)。这些标志基因的可视化显示,其空间表达模式在矢状面和冠状面切片中均保持一致(图5H)。不同切片方向中标志基因表达模式的一致性表明,SpaBatch有效校正了批次效应,实现了跨切片三维空间结构域的识别,并保持了标志基因表达的连贯性,进一步验证了其识别结果的生物学意义。最终,SpaBatch在聚类性能和批次效应校正指标上均优于基线方法(图5B和图S11B,支持信息)。
综上所述,SpaBatch通过分析小鼠脑的矢状面和冠状面数据,从宏观和微观两个层面全面描绘了成年小鼠脑的空间结构域。在矢状切片中,它有效整合了不同脑区,识别出跨切片的复杂结构,如海马体和小脑皮层;在覆盖前-后轴的35个冠状切片中,SpaBatch提供了小鼠脑区清晰的空间分布图谱,揭示了海马体和皮层等结构的动态变化,并通过各结构域特异的标志基因将两组数据关联起来。凭借其卓越的三维空间结构域识别与批次效应校正能力,SpaBatch在多项评估指标上优于其他方法,凸显其作为空间转录组分析强大工具的潜力。
利用SpaBatch识别E9.5小鼠胚胎多个组织切片间共享且相关的空间结构域
作者还将SpaBatch应用于Stereo-seq平台生成的早期小鼠胚胎数据集,旨在识别同一发育阶段(E9.5)四个切片(E2S1、E2S2、E2S3和E2S4)之间的共享空间结构域。尽管这些切片在组织结构上存在空间差异,且存在显著的批次效应,SpaBatch仍能有效将四个切片整合到嵌入空间中,并对齐其空间结构域(图6A)。作者比较了SpaBatch与STAligner、STG3Net、SEDR和SpaGIC在整合这四个连续组织切片中的表现。分析表明,SpaBatch是唯一能够在所有四个切片中完整识别出心脏区域的方法。它在空间结构域对齐方面表现出卓越能力,成功整合了全部四个切片的数据。相比之下,STAligner为最具竞争力的方法,在组织切片对齐与整合方面表现优异,但未能像SpaBatch一样在所有切片中完整捕捉心脏区域。SpaBatch不仅全面识别了小鼠胚胎的心脏区域,在其他组织的对齐方面也表现突出。它成功识别出四个连续切片中的多个共享空间结构域,包括间充质(结构域8)、生骨节(结构域9)、原始肠管(结构域10)、脑(结构域11)和脊髓(结构域12)(图S12A–C,支持信息)。SpaBatch在四个切片上的ARI、ACC和V-measure显著优于基线方法,表明其对空间结构的识别更为精确(图6D)。这种在不同空间结构域中准确捕捉和对齐多种组织结构的能力,进一步凸显了SpaBatch在处理复杂且异质的空间转录组数据方面的稳健性与通用性,有助于全面理解发育中胚胎的组织架构。
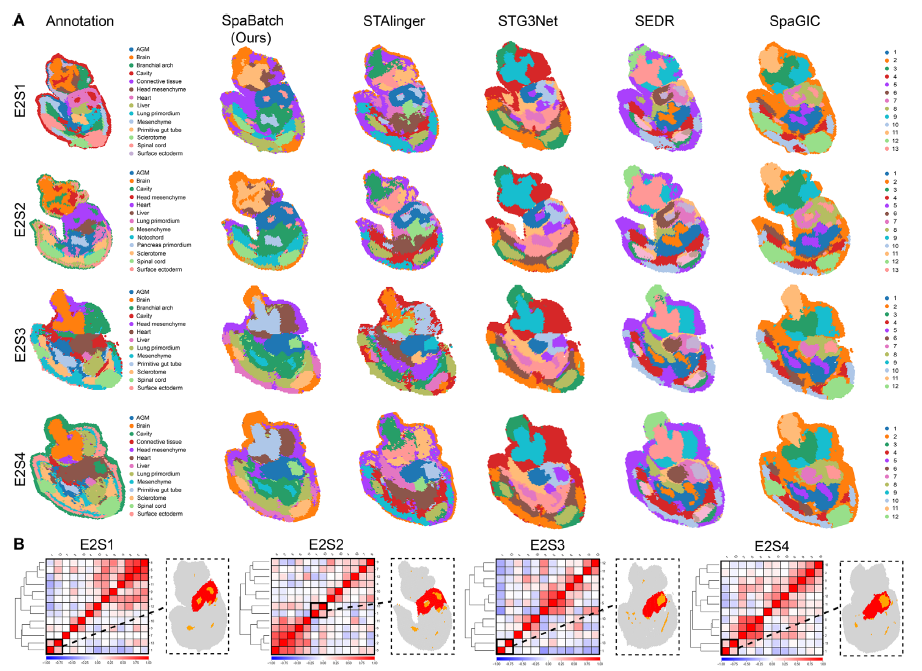
 图6 SpaBatch在小鼠胚胎数据集上跨切片匹配共享空间结构域。A)SpaBatch与其他方法在E9.5胚胎切片E2S1、E2S2、E2S3和E2S4上识别出的空间结构域。B)SpaBatch通过相关性热图识别出E9.5胚胎四个切片(E2S1、E2S2、E2S3和E2S4)中存在相关性的空间结构域1和13,这两个结构域与人工注释的心脏和腔室区域高度重叠。C)SpaBatch在全部四个切片上识别出的空间结构域(左图);SpaBatch在四个切片上识别出的心脏与腔室之间的相关区域(右图)。D)SpaBatch与其他方法在小鼠胚胎E2S1、E2S2、E2S3和E2S4切片上计算的ARI、ACC和V-measure箱线图。E)SpaBatch在四个切片上识别出与心脏和腔室相关的空间结构域(左图),以及该区域标志基因Sh3bgr的空间表达模式(右图)。
图6 SpaBatch在小鼠胚胎数据集上跨切片匹配共享空间结构域。A)SpaBatch与其他方法在E9.5胚胎切片E2S1、E2S2、E2S3和E2S4上识别出的空间结构域。B)SpaBatch通过相关性热图识别出E9.5胚胎四个切片(E2S1、E2S2、E2S3和E2S4)中存在相关性的空间结构域1和13,这两个结构域与人工注释的心脏和腔室区域高度重叠。C)SpaBatch在全部四个切片上识别出的空间结构域(左图);SpaBatch在四个切片上识别出的心脏与腔室之间的相关区域(右图)。D)SpaBatch与其他方法在小鼠胚胎E2S1、E2S2、E2S3和E2S4切片上计算的ARI、ACC和V-measure箱线图。E)SpaBatch在四个切片上识别出与心脏和腔室相关的空间结构域(左图),以及该区域标志基因Sh3bgr的空间表达模式(右图)。作者进一步通过空间结构域相关性热图探索了不同结构域之间的关系,发现结构域1与结构域13之间存在显著相关性(图6B,C)。这一发现引起作者的关注,因为这两个结构域的紧密关联可能揭示了它们在小鼠胚胎发育过程中的潜在功能联系。在后续分析中,作者将这两个结构域与胚胎中的人工注释区域进行比对,发现结构域1和结构域13与注释的心脏和腔室区域高度重叠。标志基因Sh3bgr在这四个切片的该区域均表现出显著且特异性的表达,进一步支持了二者之间的相关性(图6E)。Sh3bgr已被报道与心肌发育相关。此外,相关区域还包括结构域11和结构域12。从相关性热图中观察到,这两个空间结构域与人工注释的脑和脊髓区域高度重合(图S12A,支持信息)。作者对这两个空间结构域进行了GO功能富集分析。与脑高度相关的空间结构域主要富集于神经元发育、突触组织、轴突导向等功能,这些功能与脑组织中复杂的神经网络发育及功能调控密切相关。在与脊髓高度相关的空间结构域中,也出现了类似的神经元功能相关条目,如神经元迁移、轴突修复和运动神经元调控。这两个结构域的GO富集结果存在显著的功能重叠,表现出高度一致的功能方向(图S12D,支持信息)。这种功能上的重叠与相似性验证了相关性热图中结构域11与结构域12的高度相关性。
SpaBatch识别人类心脏的发育过程
将SpaBatch应用于人类心脏空间转录组数据集,该数据集包含在受孕后4.5-5周和6.5周(PCW)采集的两组切片(图7A),用于探究SpaBatch识别发育过程中跨切片组织结构动态变化的能力。在4.5-5 PCW阶段,SpaBatch识别出两个明显不同的空间结构域,这些结构域在6.5 PCW阶段进一步发展(图7B)。随后,作者聚焦于6.5 PCW的人类心脏,SpaBatch在九个切片中识别出六个空间结构域(图7C和图S13,支持信息)。根据先前研究提供的解剖区域注释,作者将识别出的空间结构域映射到对应的解剖区域(图7D)。例如,空间结构域1、3和4分别对应小梁心室心肌、致密心室心肌和心房心肌,这些均为心肌的重要组成部分。小梁心肌和致密心肌通常存在于心室中,其中致密心肌提供强大的收缩力,而小梁心肌则负责结构支撑和局部血流调节。心房心肌支持心脏整体的血流,并协调心房与心室之间的收缩(图7E)。通过可视化SpaBatch识别出的空间结构域比例,可直观观察到6.5 PCW人类心脏组织的动态变化。
图7 SpaBatch识别人类心脏的发育过程。A)人类心脏从受孕后4.5-5周到6.5周的发育过程。B)SpaBatch在受孕后4.5-5周人类心脏中识别出的空间结构域。C)SpaBatch在受孕后6.5周人类心脏中识别出的空间结构域。D)6.5周人类心脏的UMAP可视化及其对应的解剖区域。E)SpaBatch在6.5周人类心脏中识别出的空间结构域比例的堆叠柱状图。F)6.5周心脏中各空间结构域的GO功能富集分析。
该人类心脏数据未提供人工注释作为真实参考。针对识别出的空间结构域,作者进行了GO功能富集分析以解析其生物学功能。结构域1对应小梁心室心肌区域,其显著富集的GO条目主要与细胞呼吸相关。结构域2富集的GO条目涉及血液循环、血管形态发生和血管生成等,与其对应的解剖区域功能一致。结构域3对应致密心室心肌区域,富集过程与心脏发育和心肌收缩密切相关。结构域6的富集条目则与细胞外基质(ECM)组织相关,参与维持心血管系统的完整性与稳定性(图7F)。
SpaBatch利用有限注释实现HER2阳性乳腺癌数据集中精准的空间结构域检测
除正常组织外,作者还将SpaBatch应用于HER2阳性乳腺癌数据集,以评估其在异常组织切片中发现生物学特征的能力。该数据集包含来自8位不同患者的肿瘤样本,分为A-H共8组。其中A-D组每组包含6个切片,E-H组每组包含3个切片。作者
对A-H各组分别进行了独立分析。在A-H各组中,仅有第一个切片由病理学家进行了人工注释。作者在此数据集上对SpaBatch、STAligner、STG3Net、SEDR和SpaGIC进行了实验,并计算了各方法在第一切片上的ARI值(图S14,支持信息)。如图8A所示,SpaBatch在所有组别的全部切片中,三维空间聚类性能均优于其他方法。在整体聚类表现上,SpaBatch也取得了最高的中位数(ARI = 0.367)和均值(ARI = 0.362),显著优于排名第二的STAligner(中位数ARI = 0.196,均值ARI = 0.255),表现出明显提升(图8B)。随后作者聚焦于包含最多样空间结构域的H组数据。SpaGIC和SEDR均未能识别出原位癌区域,STG3Net未能正确识别乳腺腺体。SpaBatch与STAligner在H1切片上的表现最佳,均能准确识别出原位癌和脂肪组织等连续区域,以及乳腺腺体等离散结构。在空间结构域范围和边界识别方面,SpaBatch优于STAligner,获得了更高的ARI值(图8C)。值得注意的是,在H组的多切片联合训练中,SpaBatch仅利用H1切片的有限人工注释,即实现了跨切片的三维空间结构域识别。如H1的注释所示,SpaBatch识别出的结构域1与浸润性癌区域高度吻合,而结构域3几乎完全覆盖原位癌区域。进一步对H2和H3切片的可视化显示,预测出的结构域1和3在H&E染色图像中始终对应深色病变区域,其空间分布模式与H1切片相似。
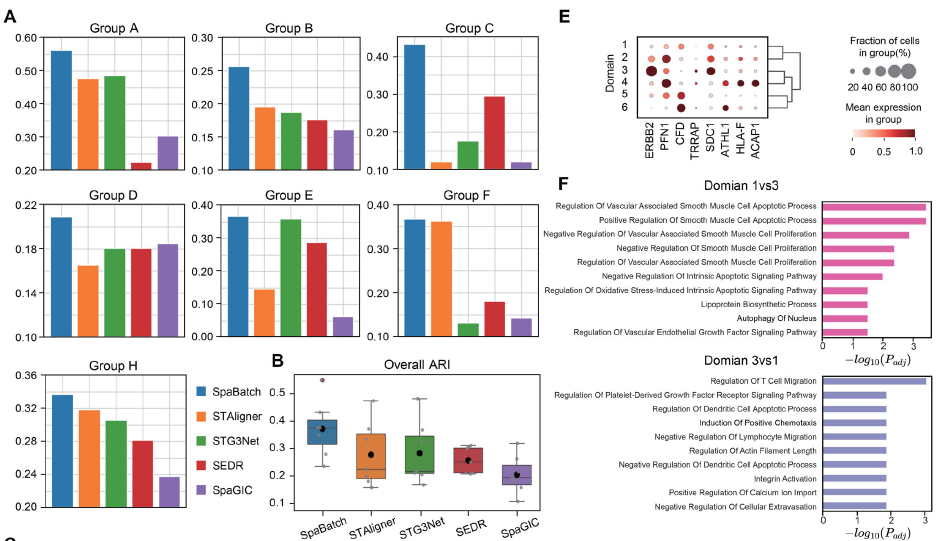
 图8 利用有限注释实现HER2阳性乳腺癌数据集中精准的空间结构域检测。A)在HER2阳性乳腺癌数据集A-H组的第一切片上,SpaBatch与其他方法相比计算得到的ARI值。B)各组ARI值的整体箱线图。C)H1切片的人工注释(左图),以及SpaBatch与其他方法在H1切片上的空间结构域识别结果(右图)。D)通过对H组数据联合训练后,SpaBatch预测出的H2和H3切片的空间结构域。E)癌组织区域与非癌区域之间差异分析的气泡图。F)原位癌与浸润性癌的GO功能富集分析结果。
图8 利用有限注释实现HER2阳性乳腺癌数据集中精准的空间结构域检测。A)在HER2阳性乳腺癌数据集A-H组的第一切片上,SpaBatch与其他方法相比计算得到的ARI值。B)各组ARI值的整体箱线图。C)H1切片的人工注释(左图),以及SpaBatch与其他方法在H1切片上的空间结构域识别结果(右图)。D)通过对H组数据联合训练后,SpaBatch预测出的H2和H3切片的空间结构域。E)癌组织区域与非癌区域之间差异分析的气泡图。F)原位癌与浸润性癌的GO功能富集分析结果。上述结果表明,SpaBatch能够有效利用有限的注释数据进行半监督学习,实现跨切片的精准空间结构域识别,而无需对每个切片都进行完整注释(图8D)。在实际研究和临床应用中,具备完整注释的空间转录组数据往往稀缺。SpaBatch仅需部分注释即可实现准确的三维空间结构域识别,从而在真实场景中推动可扩展且可靠的空間分析。作者对癌组织区域与其他区域进行了差异分析。如图8E所示,SpaBatch成功识别出从结构域3恢复出的原位癌区域相较于其他区域显著高表达乳腺癌标志基因ERBB2。随后作者进一步探究了癌组织的异质性。结构域3富集了多种与免疫反应相关的GO功能条目,包括B细胞和T细胞信号通路以及呼吸爆发调控,提示原位癌区域存在较强的免疫反应。而结构域1则富集于抑制细胞凋亡的通路,可能表明浸润性癌中存在更强的抗凋亡机制,有助于肿瘤细胞逃避免疫清除。这些生物学过程的差异凸显了两个癌区域之间的功能区别,反映了其各自独特的免疫/基质微环境特征(图8F)。
SpaBatch准确解析小鼠下丘脑视前区MERFISH平台数据并实现三维重建
为评估SpaBatch在不同测序技术中的广泛适用性,作者将该方法应用于基于成像的MERFISH技术生成的数据集,该数据集包含小鼠下丘脑的连续冠状切片。具体包括三个连续切片:下丘脑0.09、0.14和0.19,每个切片均提供人工注释。共识别出八个解剖区域,包括终纹床核(BST)、内侧视前区(MPA)、内侧视前核(MPN)、下丘脑室旁核(PV)、下丘脑室旁核(PVH)、丘脑室旁核(PVT)、第三脑室(V3)和穹窿(fx)。作者比较了SpaBatch与其他四种方法的表现,其中Scanpy代表传统分析方法。Scanpy采用主成分分析(PCA)对基因表达数据进行降维,随后进行聚类,但未引入非线性建模或明确的批次效应校正。在将聚类结果映射回原始组织切片进行可视化时,Scanpy未能捕捉到与小鼠下丘脑不同解剖结构相对应的空间模式。与其他基于深度学习的方法相比,Scanpy的聚类边界更为分散,所得聚类的空间紧凑性和一致性较低,凸显了传统方法在处理复杂空间转录组模式中的局限性。相比之下,SpaBatch在识别大多数空间结构域方面优于其他深度学习方法,且与组织学注释的一致性更高。其在三个连续冠状切片上的平均ARI得分最高(0.410),同时在ACC和V-measure指标上也表现最佳(图9A;图S15A,支持信息)。SpaBatch准确识别了BST、V3和MPA等主要区域,尤其显著的是,其生成的聚类更为紧凑,边界更平滑。相比之下,其他方法常将某一空间结构域内的点错误分配至邻近区域。
图9 SpaBatch准确解析小鼠下丘脑视前区MERFISH数据并实现三维重建。A)SpaBatch与其他方法在MERFISH平台检测的小鼠下丘脑视前区三个连续切片上识别出的空间结构域。B)SpaBatch通过校正三个连续切片的空间坐标后重建的三维空间结构域。C)全八个聚类间的差异基因表达分析,显示上调与下调基因,经调整p值小于0.01的以红色表示,大于等于0.01的以青绿色表示。D)八个聚类间差异分析的气泡图。E)SpaBatch实现空间可变成分基因Nnat在第三脑室区域(结构域6)的三维空间重建。此外,SpaBatch在批次效应去除的评估指标上也表现出更优性能(图S15A,支持信息)。为评估在SpaBatch框架内非欧几里得距离度量与欧几里得距离在MERFISH数据集上的表现差异,作者引入了基于欧几里得、曼哈顿、切比雪夫、余弦和汉明距离构建的邻接矩阵。其中,使用欧几里得距离构建的图表现最佳,其次是切比雪夫距离(图S15B,支持信息)。值得注意的是,作者引入了一种中心对齐方法,用于调整三个切片的空间坐标,从而实现二维空间结构域识别结果在三维空间中的对齐与可视化(图9B)。第三脑室(V3)是位于间脑内的中线结构,充满脑脊液,主要介于丘脑与下丘脑之间。它沿脑的正中矢状面延伸,前连侧脑室,后经中脑导水管连接第四脑室。SpaBatch准确识别出该区域,其与周围区域的边界清晰。作者进一步通过火山图对多个空间结构域进行差异分析,发现Cd24a和Nnat是空间结构域6(V3区域)的显著标志基因(图9C;图S15B,支持信息)。这些标志基因进一步通过气泡图验证其在各点上的表达比例和平均表达水平(图9D),结果确认Cd24a和Nnat在V3区域高表达,而在其他空间结构域中表达水平较低。作者对Cd24a和Nnat等标志基因进行了可视化。借助中心对齐算法对三个切片的空间坐标进行调整后,这些基因的表达模式也在三维空间中得以呈现(图9E;图S15C,支持信息)。
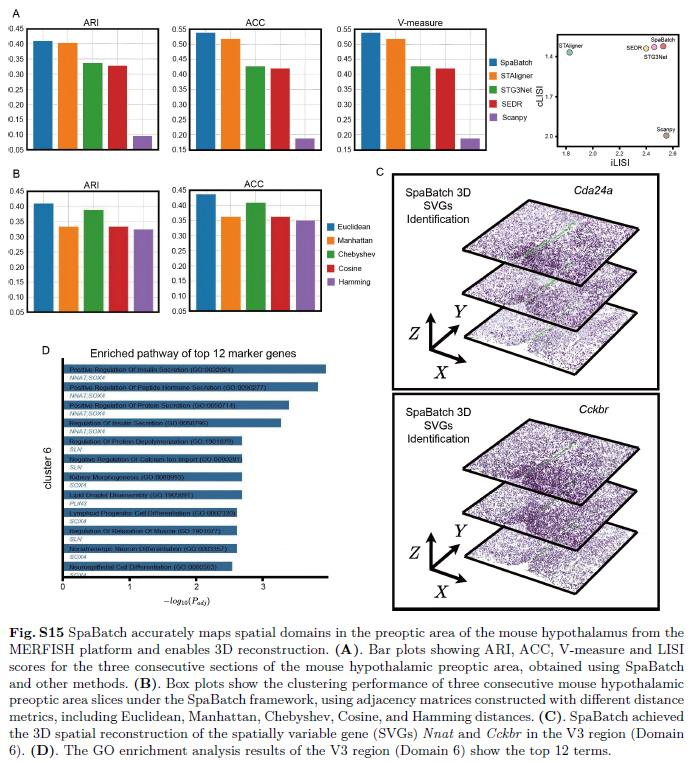
此外,作者对识别出的V3区域进行了GO功能富集分析(图S15D,支持信息)。结果揭示了多个与神经发育和分化相关的功能条目,包括去甲肾上腺素能神经元分化和神经上皮细胞分化。转录因子SOX4被识别为调控神经前体细胞分化、影响中枢和周围神经系统发育的关键因子。另一个显著富集的条目为钙离子跨膜转运的负调控,该过程与钙信号通路密切相关,而钙信号是神经元兴奋性和突触可塑性的基本调控机制。基因SLN可能通过该通路间接调控神经功能。与先前研究一致,作者还观察到葡萄糖及单糖响应通路的显著富集,这可能源于V3区域主要由室管膜细胞构成,提示其在碳水化合物转运中具有潜在作用。这些通路的富集与基因Nnat的表达密切相关。SpaBatch在MERFISH数据集上表现出色,进一步验证了其在不同技术平台、测序方法、物种和数据规模下进行三维空间结构域识别的广泛适用性与稳健性。尽管基于成像和基于测序的空间转录组技术在点阵的空间分布上存在差异,SpaBatch通过欧几里得距离构建图结构,并利用图神经网络聚合局部邻域内的特征信息,使其聚类性能不受点阵分布规则性或不规则性的影响,展现出强大的适应性与稳定性。
SpaBatch在部分重叠切片上准确识别空间结构域
为评估SpaBatch在处理部分重叠切片时的鲁棒性,作者
设计实验以检验其在整合形态差异显著的切片时的性能。在先前使用的矢状面前脑与后脑切片基础上,引入一个在结构方向和形态上均不同的冠状切片,并与两个矢状面切片进行联合整合分析(图10A)。尽管冠状面与矢状面切片在形态和空间方向上存在显著差异,但三者均来自同一组织(小鼠脑),因此具有相似的基因表达模式。SpaBatch能够有效捕捉这种跨切片的表达一致性,从而实现对结构差异较大切片的准确整合。在仅使用矢状面切片的分析中,SpaBatch成功识别出解剖学上明确的区域,如前脑的嗅球和后脑的小脑皮层。更重要的是,SpaBatch在识别跨越前脑与后脑的精细结构方面也表现出色,包括多个皮层区域和海马体(图4)。在引入空间重叠度较低的冠状切片后,SpaBatch仍保持稳健表现。模型在冠状切片中准确划分出海马体区域,其空间位置与组织学(H&E染色)图像中的对应区域高度吻合(图10B,C)。尽管相较于仅使用矢状面切片时海马体识别精度略有下降,可能源于跨切片整合过程中引入的噪声,但整体空间一致性仍保持良好。
图10 SpaBatch在部分重叠切片上准确识别空间结构域。A)小鼠前-后矢状面脑切片(来自切片1)、一个冠状面切片的H&E染色图像,以及来自艾伦脑图谱的小鼠皮层参考图谱,及其对应的具体空间亚结构域(海马体)。B)SpaBatch在小鼠脑矢状面前-后及冠状面切片上的空间结构域识别结果。C)SpaBatch在小鼠脑矢状面前-后及冠状面切片上识别出的海马体内部空间亚结构域。D)标志基因Neurod6在小鼠脑矢状面前-后及冠状面切片中的空间表达,显示其在海马体区域集中表达。E)SpaBatch识别出所有空间结构域间的相关性热图。
此外,SpaBatch在冠状切片中成功识别出位于海马体上方、具有类似层状结构的皮层区域,这些结构与矢状面切片中的皮层区域相似。在矢状面切片中识别出的结构域标志基因Neurod6,在冠状切片中的空间表达模式进一步验证了其作为海马体特异性标志基因的作用(图10D)。空间相关性矩阵也支持第10和第18聚类共同构成海马体结构,因二者在热图中相邻且具有高度相关性(图10E)。需要强调的是,部分重叠切片由于共享空间区域有限且坐标系统不一致,给跨切片对齐带来显著挑战。然而,SpaBatch仍能在该条件下实现有效的多切片整合,其技术原理如下:SpaBatch不依赖于显式的切片间邻接或配准信息,而是通过模型内共享的可训练参数,联合优化所有切片的基因表达表征,实现信息共享。虽然图结构中未构建直接的切片间连接,但块对角形式的邻接矩阵允许图神经网络在各切片内部聚合空间信息,而共享的编码器与解码器则学习一个统一的潜在空间,以捕捉跨切片的基因表达模式。该架构实现了间接的跨切片信息共享,使SpaBatch即使在切片部分重叠或形态存在差异的情况下,也能学习到一致且稳健的空间表征。因此,SpaBatch在此类场景下的有效性不仅得到实验结果的支持,也源于其模型设计的基本原理。
SpaBatch可去除基因表达噪声并增强空间标志基因的表达模式
在先前的分析中,SpaBatch识别出对应于不同皮层的标志基因,如AQP4(第1层)、CARTPT(第2层)、ENC1(第3层)、PCP4(第4层)、TMSB10(第4和第5层)以及MBP(白质)(图2)。在此基础上,作者进一步分析SpaBatch识别出的这些空间标志基因。以3号供体的四个切片为例,SpaBatch识别出的对应于皮层第1至第6层及白质的空间结构域,与大脑皮层的经典解剖分层高度一致(图S16A,支持信息)。通过可视化PCP4、TMSB10和MBP在原始数据(上图)与SpaBatch去噪后(下图)的表达空间分布,作者发现这些空间标志基因在特定皮层中强烈表达,而在其他层中表达水平较低。去噪后的表达信号更为集中和清晰,表现出更高的空间特异性,背景噪声显著降低,空间表达模式的可区分性明显增强(图S16B,支持信息)。不同皮层及其对应标志基因在原始与去噪后表达的 violin 图显示,TMSB10及其他空间标志基因(如PCP4、ENC1)在去噪后中位表达水平明显提升,分布也更为集中,反映出基因表达信号得到增强,噪声得到有效抑制(图S16C,支持信息)。红色虚线框标示出去噪效果显著的皮层与基因,进一步验证了SpaBatch通过去噪有效增强了空间结构域的生物学信号。综上所述,SpaBatch不仅能准确识别空间结构域,还可通过去噪显著改善关键标志基因的空间表达模式,有助于后续对空间生物学功能的解析与研究。
图11 SpaBatch及其消融版本(无掩码、无三元组损失、无DEC模块)在DLPFC数据集上聚类指标(ARI、ACC、V-measure)和批次校正指标(iLISI、cLISI)的比较结果。
消融实验与掩码率选择
为评估SpaBatch中各关键组件对整体性能的贡献,作者通过逐一移除模型中的三个重要模块进行消融实验,包括用于数据增强的掩码(Mask)机制、实现紧凑聚类的DEC模块以及用于批次效应校正的三元组损失(triplet loss)。图11展示了完整模型及其消融版本在DLPFC数据集上的表现,评估指标包括三项聚类指标(ARI、ACC和V-measure)以及两项批次校正指标(iLISI和cLISI)。结果表明,完整版SpaBatch模型在所有聚类指标上均达到最优或接近最优性能,说明三个模块协同作用,共同提升聚类质量。其中,移除DEC模块(w/o DEC)导致聚类性能显著下降,凸显其在引导潜在空间形成紧凑聚类结构中的关键作用。移除掩码和三元组模块同样导致聚类性能降低,表明这两个组件对提升聚类效果具有重要贡献。在批次校正方面,iLISI和cLISI结果显示,引入三元组损失的SpaBatch在数据整合与生物结构保留之间实现了最佳平衡。总体而言,掩码、DEC和三元组模块的引入对提升模型性能具有不可或缺的作用,验证了SpaBatch设计的合理性与有效性。为探究掩码率对模型性能的影响,并论证将0.2设为默认值的合理性,作者设置了从0.1到0.9、步长为0.1的掩码率进行实验。结果表明,随着掩码率增加,模型性能先上升后下降,其中0.2和0.3取得最佳效果。尽管掩码率为0.3时在箱线图中表现出更高的中位数和更紧凑的分布,但其异常值更多,导致整体均值略低于0.2,可能对下游分析引入不稳定性。因此,作者选择0.2作为默认设置,因其表现出更优的稳定性与更高的平均性能(图12)。
图12 掩码率对DLPFC数据集空间结构域识别性能的影响。掩码率设置范围为0.1至0.9,间隔0.1,分别计算A)ARI、B)ACC和C)V-measure。D)在左下角的折线图中,红线表示平均ARI值,红色阴影区域表示标准差(STD)。
为验证掩码率0.3时出现的异常值是否影响下游分析,作者在一个组织结构更复杂的小鼠脑矢状面数据集上进行了额外实验(图S17,支持信息)。当掩码率为0.2时,SpaBatch最准确地识别出相邻切片间连续的空间结构域,尤其在皮层、海马体、丘脑和下丘脑等区域表现突出。值得注意的是,仅在此设置下,海马体完整的三突触回路(包括CA1、CA2、CA3及齿状回DG)得以清晰界定。当掩码率超过0.6时,皮层区域出现不连续;而极端值如0.1和0.9则影响小脑结构的刻画。尽管如此,SpaBatch在大多数参数设置下均能稳定识别精细解剖结构,展现出良好的鲁棒性,进一步支持将0.2作为默认掩码率的选择。
不同图卷积算子的影响
GCNConv结构简单、计算效率高,能够稳定地从空间邻接图中捕捉局部表达模式,在模型稳定性、邻域结构建模能力和计算成本之间实现了良好平衡,特别适用于大规模图结构的空间转录组数据。其在多种空间转录组任务中已展现出良好有效性。为验证SpaBatch选择GCNConv作为图卷积层的合理性,作者进行了对比实验。具体地,分别将GCNConv替换为GATConv、GraphConv、SAGEConv和FeastConv,并将其集成至SpaBatch主干网络中,评估其在多切片空间结构域识别中的表现。实验在DLPFC数据集上进行,将12个组织切片按供体分为三组:供体1(151507-151510)、供体2(151669-151672)和供体3(151673-151676)。在每种图卷积配置下计算了多个聚类评估指标,包括ARI、ACC、V-measure以及LISI(iLISI和cLISI),并通过箱线图展示结果以评估性能稳定性(图S18A,支持信息)。
图S18
结果表明,GCNConv整体表现最优,在所有指标上的均值和中位数最高,方差最小,展现出较强的鲁棒性。其他卷积层如SAGEConv和FeastConv也表现尚可,ARI中位数约为0.55,说明SpaBatch对卷积类型的选择具有一定的容忍度。LISI分析进一步证实,GCNConv在缓解批次效应方面效果最佳。作者进一步可视化并展示了供体3的聚类结果(图S18B,支持信息)。GCNConv取得最高ARI得分(0.613),其次为FeastConv(0.598)和SAGEConv(0.588),而GATConv表现最差(0.448)。该差异可能源于GATConv的注意力机制对基因表达数据中的稀疏性或噪声更为敏感。与人工注释相比,除GCNConv外,其余卷积层均未能完整识别某些皮层结构。
例如,尽管SAGEConv结果接近GCNConv,但其对第6层的识别不完整,显示出性能差距。GCNConv在另外两个供体子集上也表现出色(图S19,支持信息)。作者还分析了学习所得嵌入表示的UMAP可视化结果,以评估聚类结构是否与生物学上预期的分层结构一致(图S18C,支持信息)。总体而言,GCNConv的邻域聚合策略在该数据集上表现出更优的鲁棒性,而GATConv可能需要进一步的参数调优以提升性能。需要指出的是,本实验未对不同卷积层进行架构或超参数调优,这可能带来一定不公平性,尤其是对GATConv。GATConv引入了可学习的注意力机制,能为邻近节点分配不同权重,实现更灵活自适应的特征聚合。理论上,该机制可使模型动态关注最具信息量的邻域,具备更强的表征能力和适应性。尽管GATConv在本研究中表现欠佳,可能源于其对噪声敏感或默认参数非最优,但作者认为其在空间转录组中仍具潜力。未来工作可探索引入边权重、空间位置感知或分层注意力机制等策略,以更好发挥其在复杂组织结构建模中的优势。
基于读出的三元组聚合策略中正样本数量的选择
作者引入了一种基于读出的三元组聚合策略,以应对在单个spot水平选择正样本时可能产生的误差。通过对正样本特征进行聚合后再进行对比学习,模型能够为目标结构建立更稳健和稳定的表征,从而减少因个别正样本误选所导致的错误。作者通过实验分析了在基于读出的三元组策略中,不同正样本聚合数量对模型性能的影响,并验证该策略相较于移除该模块或采用点对点对比学习的优势。具体地,作者在小鼠前脑与后脑矢状面切片数据集上,通过移除该模块(聚合正样本数=0)或逐步增加聚合正样本数量,观察SpaBatch的性能变化。如先前在DLPFC数据集上的消融实验所示,移除该模块后,所有三项聚类指标(ARI、ACC和V-measure)均明显下降。用于评估批次效应校正效果的LISI指标(包括iLISI和cLISI)也显著降低(图11)。作者发现,随着聚合正样本数量的增加,ARI得分先上升,后下降,最终趋于稳定(图S20A,支持信息)。可视化结果进一步揭示了精细结构识别的变化。
在未引入该模块时,聚类的紧凑性及空间结构域间的连续性明显受损。例如,尾状壳核(CP)区域包含大量不属于该结构域的错误分配spot,显著降低了识别准确性,同时跨切片皮层结构的对齐也变得不连贯。引入该模块后,上述问题得到显著缓解。然而,当聚合正样本数设为1时,海马体未能被完整识别,尽管较大结构域能被较准确识别。当聚合数量为2时,模型性能达到最优,成功识别出海马体等精细结构及跨切片连续区域。当数量增至4、5或6时,性能保持稳定,结果与数量为2时基本一致,但海马体腹侧部分未能被识别,整体表现略逊一筹(图S20B–D,支持信息)。随着正样本数量进一步增加,性能开始下降。作者推测,过度聚合正样本可能引入边界噪声,导致模型难以准确区分相邻但生物学上不同的空间结构域。综上所述,选择合适的正样本聚合数量对模型性能和空间结构域的准确识别至关重要。适度聚合可在稳定性与细节捕捉之间取得平衡,而过度聚合则可能导致特征混杂,性能下降。
结论
生物体和组织的生物学活动本质上是三维的。将空间转录组学局限于二维分析限制了对复杂组织结构的探索。相比单切片的空间结构域识别,多切片联合分析面临更高的技术与生物学复杂性与挑战。单切片分析仅关注单个组织切片内的空间结构,探索局部基因表达模式,适用于组织内空间异质性的初步研究。而多切片整合则分析多个连续或非连续切片的数据,不仅识别各切片内的空间结构,还需跨切片对齐与整合空间信息,构建一致的空间表达图谱。该过程必须应对切片间的批次效应与空间错位等挑战,要求算法具备强大的对齐能力与跨切片特征提取能力。随着测序技术的快速发展,越来越多高质量的空间转录组(ST)数据被产生。这迫切需要能够整合来自同一个体的多个切片、不同平台获取的同种组织切片,以及发育不同阶段的连续切片的方法。这些数据集在实验流程、测序平台、尺寸、形状和方向上往往存在差异。为应对这些挑战,作者提出SpaBatch——一种专为多切片ST数据整合与三维空间结构域识别设计的新框架。在数据预处理阶段,SpaBatch采用基于掩码的数据增强策略处理基因表达数据,以增强对稀疏性和噪声的鲁棒性。在训练过程中,SpaBatch采用预训练-微调范式:首先预训练主干VGAE网络以捕获初始空间结构与转录特征,随后引入深度嵌入聚类(DEC)模块进行微调,以增强聚类紧凑性。此外,引入基于读出策略的三元组对比损失,以显式校正批次效应。
相比STAGATE和SEDR等经典空间结构域识别方法(这些方法先利用图卷积网络学习ST数据的低维嵌入,再通过Harmony分步进行批次效应校正),SpaBatch采用端到端的联合学习框架,直接将原始输入映射到最终输出。所有中间步骤被整合进统一架构,由模型自动完成,无需后续的批次校正(补充图S21)。训练过程中,SpaBatch整合多个功能模块,同步进行嵌入学习、批次效应校正与聚类优化。这些任务通过参数共享与协同学习在单一模型内联合优化,有助于获得一致的跨切片表征与准确的空间结构域识别,同时保留底层空间结构。SpaBatch在不同数据集中均取得了大量具有生物学意义的发现。在DLPFC数据集上,SpaBatch的三维空间结构域识别结果与人工注释最为一致,准确划分出第1至第6层及白质(WM),同一皮层内的聚类混合良好。UMAP可视化进一步证实SpaBatch在批次校正上的有效性,表现出优异的切片间混合效果。在小鼠矢状面脑数据集上,SpaBatch准确识别出前脑或后脑特有的空间结构域,如嗅球和小脑皮层;更重要的是,它还能精准识别跨越前脑与后脑的共享结构域,包括背侧与腹侧海马区的连续识别,以及高度一致的皮层分层。在小鼠冠状面脑数据集上,SpaBatch进行三维空间结构域识别,并将各切片手动映射至艾伦脑图谱的小鼠皮层参考图谱,展示组织组成比例与解剖结构,从而构建了小鼠冠状面脑的综合图谱。在小鼠胚胎数据集上,SpaBatch准确识别三维空间结构域,并利用热图验证相关结构域,通过堆叠三维预测结果实现与特定解剖结构的对齐。
SpaBatch还能准确捕捉人类胚胎心脏的动态变化。在HER2阳性乳腺癌数据集中,SpaBatch利用首张切片的有限人工注释进行半监督学习,引导训练过程,实现其余切片的准确空间结构域识别,并精确标注原位癌、浸润性癌等区域。在多种空间转录组数据集中,SpaBatch展现出卓越的性能与广泛的适用性,不仅实现高精度的三维空间结构域识别,还揭示了具有生物学意义的结构。在使用MERFISH技术测序的小鼠下丘脑切片中,SpaBatch准确识别出第三脑室(V3)细长而精细的解剖结构,并通过中心对齐算法在三维空间重建其结构域与空间标志基因。综上所述,SpaBatch是一种通用且高效用于三维空间结构域识别的框架,能有效应对多切片ST数据在来源、尺度、方向和批次效应上的异质性。通过整合基于掩码的增强、预训练-微调策略、深度聚类与三元组对比学习,SpaBatch显著提升了聚类的准确性与一致性,同时实现了跨切片对齐与批次可迁移性。在多种实际数据集中,SpaBatch成功重建复杂组织架构,揭示关键生物学特征与发育动态,在结构分析、发育生物学与疾病表征中展现出巨大潜力。SpaBatch为推动空间转录组学从二维走向三维、从静态走向动态提供了强有力的技术支持,在未来研究与应用中前景广阔。
总结-概述-分析方法
本篇研究提出了一种名为 SpaBatch 的深度学习框架,用于空间转录组学(ST)数据的跨切片整合和 3D 空间域识别。该方法旨在解决现有方法主要关注 2D 空间域识别、未能充分考虑切片间空间相关性和批次效应校正的限制。以下是对 SpaBatch 框架的实验设计、分析方法和结果内容的全面总结。
实验设计与分析方法
1. 数据集和预处理
研究使用了八个真实世界的 ST 数据集进行验证,这些数据集涵盖了不同的组织类型(如人皮层、小鼠大脑、人胚胎心脏、HER2+ 乳腺癌)、物种和技术平台(如 10x Visium、Stereo-seq、MERFISH)。
预处理步骤: 输入数据包括多切片的基因表达矩阵和空间位置。首先,沿斑点维度连接基因表达矩阵。删除低表达的基因,随后进行对数转换和基于文库大小的标准化。最后,选择 Top-2000 个高变基因(HVGs),并应用主成分分析(PCA)将维度降低至 $X \in R^{n \times m}$。
2. SpaBatch 模型架构与分析方法
SpaBatch 框架的核心是一个两阶段策略(预训练和微调),并集成了三种关键技术模块。
A. 核心组件和数据增强:
掩码机制(Mask Mechanism): 在数据预处理阶段引入,旨在增强模型对 ST 数据中常见稀疏性和技术噪声的鲁棒性。该策略随机掩盖一部分斑点,并用可学习的嵌入向量替换其基因表达向量。默认掩码率 $\rho$ 设为 0.2。
空间图构建(Spatial Graph Construction): 通过计算斑点间的欧氏距离,为每个切片独立选择 $k$ 近邻($k$ 在 6 到 12 之间,默认设置为 8)来构建空间图。随后,各切片的邻接矩阵以块对角形式连接起来,表示为 $A \in R^{n \times n}$。
潜空间表征学习(Latent Representation Learning):
骨干网络 (VGAE): 在预训练阶段,使用掩码变分图自编码器(VGAE)共同建模基因表达和空间邻接信息,学习低维潜嵌入 $Z$。VGAE 编码器包含全连接层和图卷积网络(GCN)层。解码器部分使用内积解码器重建邻接矩阵 $\tilde{A}$,并使用 GCN 层重建原始基因表达矩阵 $\tilde{X}$。
VGAE 损失函数 $\mathcal{L}_{\text{VGAE}}$ 包括:图重建损失 $\mathcal{L}_{\text{graph}}$、KL 散度损失 $\mathcal{L}_{\text{KL}}$(用于潜空间正则化)、以及基于掩码自监督的 Scaled Cosine Embedding (SCE) 损失 $\mathcal{L}_{\text{sce}}$。
B. 批次效应校正与聚类优化:
自监督模块(DEC): 在微调阶段引入深度嵌入式聚类(DEC)模块,通过迭代优化聚类中心,增强已识别域内的空间紧凑性。该模块通过最小化目标分布 $p_{ij}$ 和软分配概率 $q_{ij}$ 之间的 KL 散度($\mathcal{L}_{\text{DEC}}$)来实现。
基于读出聚合的 Triplet 对比学习: 引入该机制以解决切片间变异性和减轻批次效应。它首先基于低维嵌入识别跨切片的互最近邻(MNNs)作为锚点。不同于依赖单个 MNN 对,SpaBatch 采用读出聚合策略,将锚点的 $\alpha$ 个最相似邻居聚合成单个正样本表示 $z^+_i$,并聚合 $\alpha$ 个随机采样的不相似斑点为负样本表示 $z^-_i$。Triplet 损失 $\mathcal{L}_{\text{Tri}}$ 用于最小化锚点-正样本距离,同时最大化锚点-负样本距离。
总体损失函数: 最终的整体损失函数 $\mathcal{L}_{\text{overall}}$ 是 $\mathcal{L}_{\text{VGAE}}$、$\mathcal{L}_{\text{DEC}}$ 和 $\mathcal{L}_{\text{Tri}}$ 三者的加权组合。
C. 3D 空间聚类与评估标准:
聚类方法: 从 SpaBatch 学习到的低维潜表征随后使用 R 语言中的 mclust 包进行聚类,以识别空间域。
聚类评估指标: 使用 Adjusted Rand Index (ARI)、Average Clustering Consistency (ACC)(结合 NMI 和 AMI)、以及 V-measure(结合同质性 HOM 和完整性 COM) 来评估聚类结果与手动注释的一致性。
批次效应评估指标: 使用 Integration Local Inverse Simpson’s Index (iLISI) 来评估不同批次(切片)在局部邻域内的混合程度(iLISI 越高,批次混合越好);使用 Cell-type Local Inverse Simpson’s Index (cLISI) 来评估空间域的分离度(cLISI 越低,域分离越好)。SpaBatch 旨在实现 iLISI 和 cLISI 的平衡。
结果内容总结
SpaBatch 在所有测试数据集上持续优于先进的方法。
1. 人 DLPFC 数据集
3D 空间域识别: SpaBatch 在空间域识别方面表现出最高的聚类准确性,在 Sample 3 上平均 ARI 达到 0.613,优于所有其他方法。
批次效应校正: SpaBatch 在 iLISI 和 cLISI 上实现了平衡性能,表明它在有效校正批次效应的同时,保留了重要的生物学信号。UMAP 可视化结果显示,SpaBatch 实现了不同批次间的平滑和均匀混合。
组织结构: 准确捕获了皮层的保守组织结构,包括 L1 至 L6 层和白质(WM)。
2. 小鼠大脑数据集(宏观与微观)
Sagittal 切片(宏观): SpaBatch 能够准确识别跨区域的共享结构,例如嗅球和小脑皮层。在更精细的组织结构上,SpaBatch 成功识别了海马腹侧区域,并完整识别了由 CA1, CA2, CA3 和齿状回(DG)组成的经典三突触回路。在 Section 1 的前部切片上,SpaBatch 达到了最高的 ARI (0.44)。
Coronal 切片(微观): SpaBatch 分析了跨前-后轴的 35 个冠状切片,揭示了小鼠大脑区域的动态变化,例如海马区在切片 15A 出现,在切片 31A 后消失。它甚至能将皮层细分为手动注释中未区分的区域(如 FRP 1 和 FRP 2/3)。
Marker 基因: 跨不同切片方向(Sagittal 和 Coronal)观察到标记基因(如 $Neurod6, Lamp5, Ncald$)的表达模式保持一致,证明了 SpaBatch 整合的连贯性。
3. 早期小鼠胚胎数据集
SpaBatch 有效整合了 E9.5 阶段的四个切片。它是唯一能够完整识别所有四个切片中的心脏区域的方法。
通过空间域相关性热图,SpaBatch 发现了域 1 和域 13 之间的高度相关性,它们与手动注释的心脏和腔区域高度重叠,并由心脏发育相关标记基因 $Sh3bgr$ 表达所支持。
4. 人胚胎心脏发育数据集
SpaBatch 捕获了人胚胎心脏从 4.5-5 PCW 到 6.5 PCW 的动态时空变化。
在 6.5 PCW,SpaBatch 识别了六个空间域,并将其映射到解剖区域,如小梁室心肌、致密室心肌和心房心肌。GO 富集分析验证了这些区域的功能(如细胞呼吸、心脏发育)。
5. HER2+ 乳腺癌数据集
半监督学习: SpaBatch 仅利用第一张切片的有限注释,以半监督方式指导训练,成功在其他未注释切片中准确识别了空间域,如原位癌和浸润性癌区域。
聚类性能: 总体聚类性能最高,中位 ARI 达到 0.367,显著优于第二名 STAligner。
生物发现: 识别出原位癌区域高表达乳腺癌标记物 $ERBB2$。原位癌区域富集了免疫反应相关的 GO 术语,而浸润性癌区域富集了抑制细胞凋亡的通路。
6. MERFISH 小鼠下丘脑数据集
SpaBatch 适用于基于成像的技术平台(MERFISH),并达到了最高的平均 ARI 0.410。
它准确识别了第三脑室 (V3) 等细长、精细的解剖结构。通过中心对齐算法,SpaBatch 实现了空间域和空间标记基因(如 $Nnat$)的 3D 重建。
7. 鲁棒性与消融实验
部分重叠切片: SpaBatch 在处理形态学差异显著且空间重叠度低的切片(如 Sagittal 与 Coronal 小鼠脑切片)时表现稳健。这得益于其不依赖显式切片间对齐或配准,而是通过共享参数在统一潜空间中学习基因表达模式。
基因表达去噪: SpaBatch 能显著改善关键标记基因(如 $PCP4, TMSB10$)的空间表达模式,增强信号特异性并降低背景噪声。
消融实验: 结果证实,掩码机制、DEC 模块和Triplet 损失这三个核心组件均不可或缺,它们协同作用增强了聚类质量,并在数据整合和生物结构保留方面取得了最佳平衡。默认掩码率 0.2 被证明是最稳定且平均性能最高的设置。
本研究中开发的新生物计算方法是 SpaBatch 框架,它是一个用于空间转录组学(ST)数据的多切片整合和 3D 空间域识别的深度学习框架。SpaBatch 采用了两阶段训练策略(预训练和微调)以及多种创新模块,旨在解决现有方法中切片间空间关联性不足和批次效应校正不稳定等挑战。以下是对 SpaBatch 框架中使用的分析计算方法、模型优化参数和公式的详细总结:
数据预处理与增强
SpaBatch 接收多切片的基因表达矩阵和空间位置作为输入。
1. 标准化和降维
在构建多切片基因表达矩阵后,移除低表达基因(在少于 50 个细胞中表达且总表达计数小于 10 的基因)。随后,使用 SCANPY 包进行对数转换和基于文库大小的标准化。最后,选择 Top-2000 个高变基因,并应用主成分分析(PCA)将维度降低,得到输入矩阵 $X \in R^{n \times m}$。
2. 掩码机制 (Mask Mechanism)
为了增强模型对 ST 数据中常见的高稀疏性和技术噪声的鲁棒性,SpaBatch 在数据预处理阶段引入了基于掩码的数据增强策略。该策略随机掩盖一部分斑点,并将其基因表达向量替换为可学习的嵌入向量 $x^{[\text{mask}]}$。
掩码计算公式 (1):
$$\tilde{x}_i = \begin{cases} x^{[\text{mask}]}, & \text{if } V_i \in V_m \\ x_i, & \text{if } V_i \notin V_m \end{cases} \quad (1)$$
其中 $V_i$ 是第 $i$ 个斑点,$\tilde{x}_i$ 是掩码后的特征向量,$V_m$ 是被随机采样的掩码斑点集合。
默认掩码率 $\rho$:0.2。
3. 空间图构建 (Spatial Graph Construction)
通过计算斑点间的欧氏距离,为每个斑点选择 $k$ 近邻来构建空间图。
$k$ 值在 6 到 12 之间可调,默认设置为 8。
构建的每个切片的邻接矩阵 $A$ 以块对角形式连接起来,表示为 $A \in R^{n \times n}$。
潜空间表征学习
SpaBatch 使用两阶段策略:预训练阶段主要学习潜在嵌入,微调阶段引入自监督模块和对比学习进行聚类优化和批次校正。
1. 预训练阶段:掩码变分图自编码器 (VGAE)
VGAE 是 SpaBatch 的骨干网络,由编码器和解码器组成,共同建模基因表达和空间邻接信息。
编码器 (Encoder):
特征提取: 两个全连接层将 $X^{[\text{mask}]}$ 转化为低维表示 $Z_f \in R^{n \times d_f}$。
图卷积网络 (GCN): 用于整合空间信息。第一层 GCN 学习共享表示 $Z'_g$ (公式 2),第二层 GCN 独立建模均值 $\mu$ 和方差 $\log(\sigma^2)$ (公式 3)。
GCN 公式 (2):
$$Z'_g = \text{GCN}(A, Z_f) = \text{ReLU}(\text{BN}(\hat{A} Z_f W_0))$$
其中 $\hat{A} = D^{-1/2} A D^{-1/2}$,BN 代表 Batch Normalization。
均值 $\mu$ 和方差 $\log(\sigma^2)$ 公式 (3):
$$\mu = \text{GCN}(A, Z'_g) = \text{ReLU}(\text{BN}(\hat{A} Z'_g W_\mu))$$
$$\log(\sigma^2) = \text{GCN}(A, Z'_g) = \text{ReLU}(\text{BN}(\hat{A} Z'_g W_\sigma))$$
重参数化技巧 (4): 用于构造最终的潜在表示 $Z_g$。
$$Z_g = \mu + \log(\sigma^2) * \epsilon \quad (4)$$
最终潜在嵌入 $Z = Z_f + Z_g$。
编码器网络维度: 全连接层维度为 64 和 16;图卷积层维度为 64 和 16。
解码器 (Decoder):
邻接矩阵重建 ($\tilde{A}$): 使用内积解码器计算节点对之间存在边的概率。
公式 (6):
$$\tilde{A} = p(A_{ij} = 1 | z_i, z_j) = \text{sigmoid}(Z Z^T) \quad (6)$$
图重建损失 $\mathcal{L}_{\text{graph}}$ (7):
$$\mathcal{L}_{\text{graph}} = ||A - \tilde{A}||^2 \quad (7)$$
基因表达重建 ($\tilde{X}$): 使用单层 GCN 从 $Z$ 重建原始基因表达矩阵。
Scaled Cosine Embedding (SCE) 损失 $\mathcal{L}_{\text{sce}}$ (9): 用于最小化 $X^{[\text{mask}]}$ 和 $\tilde{X}^{[\text{mask}]}$ 之间的差异。
$$\mathcal{L}_{\text{sce}} = \frac{1}{|\mathcal{V}_m|} \sum_{v_i \in \mathcal{V}_m} \left(1 - \frac{x^{[\text{mask}]}_i \tilde{x}^{\text{T}[\text{mask}]}_i}{\lVert x^{[\text{mask}]}_i \rVert \lVert \tilde{x}^{[\text{mask}]}_i \rVert}\right)^\gamma, \quad \gamma \geq 1 \quad (9)$$
$\gamma$ 是用于调整模型对大误差敏感度的缩放因子,$\gamma \geq 1$。
KL 散度损失 $\mathcal{L}_{\text{KL}}$ (8): 鼓励潜在空间匹配先验分布(标准正态分布)。
总体 VGAE 损失 $\mathcal{L}_{\text{VGAE}}$ (10):
$$\mathcal{L}_{\text{VGAE}} = \mathcal{L}_{\text{graph}} + \mathcal{L}_{\text{KL}} + \mathcal{L}_{\text{sce}} \quad (10)$$
2. 微调阶段:聚类和批次校正
微调阶段引入了深度嵌入式聚类(DEC)和基于读出聚合的 Triplet 对比学习模块。
A. 深度嵌入式聚类 (DEC) 模块
用于迭代优化聚类中心,增强已识别域内的空间紧凑性。
软分配概率 $q_{ij}$ (11): 使用 Student’s t-分布相似度量化斑点 $i$ 属于聚类中心 $\phi_j$ 的关系。
$$\text{q}_{ij} = \frac{(1 + \lVert z_i - \phi_j \rVert^2)^{-1}}{\sum_{j'} (1 + \lVert z_i - \phi_{j'} \rVert^2)^{-1}} \quad (11)$$
目标分布 $p_{ij}$ (12): 通过增强当前软分配分布的峰值来提高模型区分不同聚类的能力。
$$\text{p}_{ij} = \frac{q^2_{ij} / \sum_i q_{ij}}{\sum_{j'} (q^2_{ij'} / \sum_i q_{ij'})} \quad (12)$$
DEC 损失 $\mathcal{L}_{\text{DEC}}$ (KL 散度) (13): 最小化目标分布 $P$ 和软分配概率 $Q$ 之间的 KL 散度。
$$\mathcal{L}_{\text{DEC}} = \text{KL}(P|Q) = \sum_i \sum_j p_{ij} \log \frac{p_{ij}}{q_{ij}} \quad (13)$$
聚类中心数量 $J$ (默认): 20。
B. 基于读出聚合的 Triplet 对比学习 (Triplet Contrastive Learning)
用于解决跨切片批次效应和变异性问题。它通过聚合多个正样本来表示局部微环境,以增强模型对空间噪声的鲁棒性。
锚点选择: 基于低维嵌入,识别跨切片互最近邻(MNNs)作为锚点 $z_i$。
读出聚合策略:
选择 $\alpha$ 个最相似邻居 $z \in \mathcal{V}^+_i$ 聚合为正样本表示 $z^+_i = \frac{1}{\alpha} \sum_{z \in \mathcal{V}^+_i} z$。
随机采样 $\alpha$ 个不相似斑点 $z \in \mathcal{V}^-_i$ 聚合为负样本表示 $z^-_i = \frac{1}{\alpha} \sum_{z \in \mathcal{V}^-_i} z$。
Triplet 损失 $\mathcal{L}_{\text{Tri}}$ (14): 最小化锚点-正样本距离,同时最大化锚点-负样本距离。
$$\mathcal{L}_{\text{Tri}} = \frac{1}{N_{\text{Tri}}} \sum_{i=1}^{N_{\text{Tri}}} \max (\lVert z_i - z^+_i \rVert^2 - \lVert z_i - z^-_i \rVert^2 + \tau, 0) \quad (14)$$
其中 $N_{\text{Tri}}$ 是 Triplet 集合的数量,$\tau$ 是间隔(默认值为 1.0)。
3. 总体优化和损失函数
SpaBatch 采用端到端的联合学习框架。在训练阶段,模型优化总体损失函数 $\mathcal{L}_{\text{overall}}$。
总体损失函数 (15):
$$\mathcal{L}_{\text{overall}} = \lambda_1 \mathcal{L}_{\text{VGAE}} + \lambda_2 \mathcal{L}_{\text{DEC}} + \lambda_3 \mathcal{L}_{\text{Tri}} \quad (15)$$
其中 $\lambda_1, \lambda_2, \lambda_3$ 是用于平衡三个损失项贡献的系数。
更新频率: $\mathcal{L}_{\text{VGAE}}$ 持续优化;$\mathcal{L}_{\text{DEC}}$ 每 20 个 epoch 更新一次;$\mathcal{L}_{\text{Tri}}$ 每 500 个 epoch 更新一次。
优化器与参数: 使用 Adam 优化器,学习率设置为 $5e-4$,权重衰减设置为 $1e-4$。
3D 空间聚类与评估方法
1. 空间聚类方法
从 SpaBatch 学习到的低维潜在表征 $Z$ 随后使用 R 语言中的 mclust 包 (v6.0.1) 进行聚类,以识别空间域。
2. 聚类评估指标
Adjusted Rand Index (ARI): 用于衡量聚类结果与手动注释的相似度。
$$\text{ARI} = \frac{TP + TN - E}{TP + TN + FP + FN - E} \quad (16)$$
Average Clustering Consistency (ACC): 结合了 Normalized Mutual Information (NMI) 和 Adjusted Mutual Information (AMI)。
$$\text{ACC} = \frac{\text{NMI} + \text{AMI}}{2} \quad (18)$$
V-measure: 基于信息论,是同质性 (Homogeneity, HOM) 和完整性 (Completeness, COM) 的调和平均数。
$$\text{V-measure} = \frac{2 \cdot \text{HOM} \cdot \text{COM}}{\text{HOM} + \text{COM}} \quad (19)$$
3. 批次效应评估指标
Integration Local Inverse Simpson’s Index (iLISI): 评估批次效应去除的有效性。iLISI 越高,表明不同批次(切片)在局部邻域内的混合度越好。
Cell-type Local Inverse Simpson’s Index (cLISI): 评估空间域分离度。cLISI 越低,表明空间域(或细胞类型)分离越好。SpaBatch 旨在实现 iLISI 和 cLISI 的平衡性能。
论文数据分析复现(用户指南)
conda create -n spabatch_env python==3.11conda activate spabatch_env
pip install -r requirements.txt
文献原文,附件和新分析方法的复现(包含分析数据和梳理好的代码)可以后台获取下载链接,关键词:250930(后台发送信息输入关键词250930,自动回复下载链接);针对研究论文中的分析,任何问题都可以留言或私信询问。
我们也创建了一个交流群,平时大家可以一起学习交流,我们也会花时间维护(当前群人数满了,有兴趣进群的可以后台私信,我们拉你进群交流):
团队信息
Min
Lab
https://wenwenmin.github.io/
机器学习在计算生物学中的应用 || PART I || 2025 (点击扫码观看)

![]()
![]()
机器学习在计算生物学中的应用 || PART II || 2025(点击扫码观看)

![]()
![]()
空间组学的实用分析方法 || ISB-线上培训 (点击扫码观看)

![]()
![]()
通过计算阵列重建的可扩展空间转录组学 || The Chen Lab(点击扫码观看)
空间转录组学的模型和方法 || Ben Raphael || CGSI(点击扫码观看)

![]()
![]()
更多资源欢迎关注B站(关注MCBRLab )
其他参考基础模型:
Nature Methods || 大规模单细胞转录组学基础模型 || scFoundation
scGPT-spatial:面向空间转录组学的单细胞基础模型(scGPT || Nature Methods)的持续预训练
Nature || 2024 HCA || SCimilarity:一种用于大规模搜索相似人类细胞的细胞图谱基础模型-单细胞注释
Nature || 2024 HCA || 人类神经类器官的综合转录组细胞图谱-单细胞注释
Nature Genetics || 2024 || 人类乳腺细胞图谱 || 单细胞图谱能够映射成人人体乳腺的稳态细胞变化
综述:利用最优传输技术分析单细胞和空间组学数据 || Nature Reviews Methods Primers
Nature Genetics || 利用高级统计方法(潜在嵌入多元回归)解析多条件下的单细胞组学数据
Nature Methods || 综述:单细胞多组学中的小样本方法:单个数据点的重要性
Nature Reviews Genetics || 综述:单细胞多组学时代的基因调控网络推断
参考文献
Niu J, Fang D, Chen J, Xiong Y, Liu J, Min W. SpaBatch: Deep Learning-Based Cross-Slice Integration and 3D Spatial Domain Identification in Spatial Transcriptomics. Adv Sci (Weinh). Published online September 15, 2025. doi:10.1002/advs.202509090









